Согласно последним данным [1], толстый кишечник человека населен приблизительно 100 трлн бактерий, относящихся к 36 000 видов (1011 бактерий на грамм кишечного содержимого), образующим его микрофлору. Считается, что количество генов представителей кишечной микрофлоры превышает аналогичное у человека более чем в 150 раз [2], что в сочетании с открытием у данных организмов ряда жизненно необходимых функций позволило сделать вывод о Homo sapiens как о «суперорганизме», коэволюционно развивающемся параллельно со своей микрофлорой [3]. Геном такого «суперорганизма», таким образом, целесообразно представлять как «метагеном», включающий в себя гены собственно человека как биологического вида и гены организмов, составляющих его микрофлору [3]. Основная сложность в детальном определении состава кишечной микрофлоры до последних лет состояла в том, что около 70–80% ее представителей не культивируются в лабораторных условиях [4]. Вместе с тем последние достижения в области методов молекулярной биологии позволили сделать очередной прорыв в микробиологии и приступить к интенсивной расшифровке качественного и количественного состава микрофлоры кишечника, а также к детальному изучению взаимодействий составляющих ее организмов как с хозяином, так и между собой [4]. Фундаментальные сравнительные исследования привели к пониманию того факта, что каждый человек обладает уникальным составом микрофлоры, который во многом определяется в течение первых лет его жизни, однако подвержен значительным обратимым и необратимым изменениям под влиянием множества факторов (возраст, изменение условий окружающей среды, тип питания, применение различных лекарственных средств, патологические процессы в организме и др.) [5]. Абсолютное большинство представителей микрофлоры кишечника являются бактериями, более 99% из них относятся к 7 основным типам: Firmicutes, Bacteroidetes (эти два типа являются доминирующими), Proteobacteria, Actinobacteria, Fusobacteria, Verrucomicrobiaи Cyanobacteria (так называемые субдоминантные типы) [6]. Эти микроорганизмы играют важную роль в поддержании как местного, так и системного гомеостаза хозяина-макроорганизма, синтезируя ряд витаминов и аминокислот, ферментируя непереваренные субстраты и эндогенную слизь (что приводит к образованию короткоцепочечных жирных кислот, являющихся источником энергии и стимулирующих всасывание солей и воды), принимая участие в биотрансформации желчных кислот (что существенно для метаболизма холестерола и глюкозы), способствуя анатомическому развитию кишечника, развитию и регуляции работы иммунной системы (особенно местной), препятствуя колонизации толстого кишечника патогенными агентами, стимулируя выработку слизи и антимикробных белков, укрепляя барьер между полостью и тканями кишечника, регулируя витальные процессы клеток кишечника (пролиферацию, рост, дифференцировку, выживание и гибель), контролируя ангиогенез в окружающих тканях и выполняя ряд других, менее необходимых функций [7]. Таким образом, при нарушениях функционирования кишечной микрофлоры повышается риск развития значительного числа заболеваний (инфекционных, метаболических, аутоиммунных, онкологических, неврологических и эндокринных) [4]. В данном обзоре анализируется роль микрофлоры кишечника в развитии колоректального рака. Особую актуальность этой проблеме придает то, что данный тип злокачественных опухолей является третьим по распространенности и четвертым по смертности в мире [8]. При выявлении заболевания на поздней стадии 5-летняя выживаемость не превышает 10%, однако при обнаружении на ранней стадии этот показатель может достигать 90% [8]. Следовательно, проблема разработки и внедрения в практику новых методов определения риска развития колоректального рака для профилактики и ранней диагностики стоит достаточно остро, при этом новые предиктивные маркеры вероятности возникновения этого типа злокачественных новообразований вполне могут быть найдены в изменениях качественного и количественного состава кишечной микрофлоры [9].
Механизмы канцерогенеза, ассоциированные с микрофлорой кишечника человека
1. Метаболические механизмы
Основой действия данной группы механизмов является то, что представители кишечной микрофлоры принимают активное участие в деградации, модификации, биотрансформации и детоксификации химических соединений, поступающих в толстый кишечник после приема пищи и ее переработки в более проксимальных отделах желудочно-кишечного тракта.
Так, S.J. O’Keefe и соавт. [10] обратили внимание на то, что колоректальный рак среди коренного населения Африки встречается в 65 раз реже, чем среди афроамериканцев. В своем исследовании эти авторы обнаружили, что такая зависимость заболеваемости от региона проживания независимо связана, помимо остальных факторов, с обильным употреблением мяса и животных жиров, что ведет к увеличению содержания в кишечнике афроамериканцев 7α-дегидроксилирующих бактерий (в частности, рода Clostridium). Известно, что данные микроорганизмы трансформируют первичные желчные кислоты (холевую и хенодезоксихолевую) до вторичных (холевой и литохолевой), обладающих цитотоксическим и мутагенным, и, следовательно, канцерогенным действием [11]. Одним из способов детоксификации желчных кислот и их ускоренного выведения из организма является их сульфатирование, поскольку в результате этой модификации они становятся более водорастворимыми и менее подвержены кишечно-печеночной рециркуляции [12]. В исследовании A. Roessner и соавт. показано, что некоторые представители родов Clostridium, Bacteroidesи Bifidobacteriumобразуют арилсульфатазу, десульфатирующую преобразованные желчные кислоты и поэтому способствующую канцерогенезу [13]. Кроме того, мясо содержит много серосодержащих аминокислот и поэтому является богатым источником пищевой серы, стимулирующей рост сульфатредуцирующих бактерий (Desulfovibriospp., Desulfobacterspp., Desulfobulbusspp., Desulfomonasspp., Desulfotomaculumspp.), образующих в результате своей деятельности сероводород [12]. Это химическое соединение способствует образованию свободных радикалов, угнетает утилизацию являющегося антиканцерогеном бутирата, ингибирует продукцию слизи и метилирование ДНК. Кроме того, нарушение его детоксификации также может приводить к развитию колоректального рака [14].
A.J. McBain и соавт. [13] выявили, что населяющие кишечник бактерии в большом количестве продуцируют гликозидгидролазы (глюкозидазы), действие которых приводит к появлению в кишечнике агликонов, некоторые из которых (агликон цикозина) обладают цитотоксическим, тератогенным и радиомиметическим действием. Ряд содержащихся в пище химических соединений метаболизируется в печени, подвергается конъюгации с глюкуроновой кислотой и по желчным протокам попадает в кишечник, где расщепляется до канцерогенов образующими β-глюкуронидазу бактериями (часть представителей рода Clostridium) [15], причем в фекалиях больных колоректальным раком активность этого фермента выше, чем в содержимом кишечника пациентов контрольной группы [16]. По аналогии с β-глюкуронидазой вырабатываемая многими бактериями (в особенности Clostridiumclostridioforme, Enterococcusfaecalis, Ruminococcusobeum, Bifidobacteriumadolescentis) азоредуктаза метаболизирует водорастворимые азокрасители и некоторые химиопрепараты до канцерогенных ароматических аминов [17]. Более того, ароматические амины также могут образовываться из ароматических нитросоединений под воздействием синтезирующихся в кишечнике бактериальных нитроредуктаз [18]. Алкогольдегидрогеназа, преобразующая этанол в канцерогенный ацетальдегид, также может продуцироваться бактериями микрофлоры кишечника, поскольку уровень ацетальдегида в слизистой оболочке кишечника гнотобионтов был существенно ниже, чем у традиционно используемых лабораторных грызунов [19], и кодирующая этот фермент нуклеотидная последовательность была найдена в большинстве известных геномов микробов, населяющих желудочно-кишечный тракт [20]. Бактерии, относящиеся к типу Bacteroidetes, образуют фекапентены, нарушающие работу систем антиоксидантной защиты путем окисления глутатиона и вызывающие мутации [21]. Кроме того, одним из основных метаболических механизмов канцерогенеза является индукция представителями микрофлоры кишечника образования активных форм кислорода, которые обладают прямым мутагенным эффектом, влияя на все основные витальные процессы в клетках кишечного эпителия [22, 23].
Отдельно стоит выделить роль микрофлоры кишечника в ассоциированном с ожирением канцерогенезе. Так, F. Backhed и соавт. [24] показали, что количество жира в организме гнотобионтов после пересадки им микрофлоры кишечника традиционных лабораторных мышей увеличивалось на 60% в течение 2 нед, и у них также значительно повышалась резистентность к инсулину. Более того, колонизация гнотобионтов микрофлорой, предположительно ассоциированной с ожирением, приводила к бoльшему набору массы жира в организме, чем в случае их колонизации иной микрофлорой [24]. Эти результаты позволили предположить, что особенности микрофлоры кишечника являются дополнительным фактором, играющим важную роль в развитии ожирения.
2. Иммунные механизмы
Хроническое воспаление существенно повышает вероятность неопластической трансформации клеток окружающих тканей, индуцируя мутации и эпигенетические перестройки через образование свободных радикалов, ингибируя апоптоз, способствуя снижению клеточной дифференцировки, а также стимулируя клеточную пролиферацию, перестройку стромы, процессы ангиогенеза и инвазии [25]. Аномальные воспалительные процессы в толстом кишечнике не только препятствуют существованию нормальной микрофлоры, но и способствуют колонизации этого органа патогенными микроорганизмами, отсутствующими в норме и вызывающими на себя иммунный ответ, в результате чего «порочный круг воспаления» замыкается [26]. Кроме того, чужеродные патогенные микробы способны индуцировать развитие воспалительного процесса и без предшествующих нарушений гомеостаза в тканях кишечника, изменяя тем самым постоянное состояние физиологического воспаления, вызываемое нормальной микрофлорой, на состояние патологического воспаления [27]. Основные механизмы таких патологических воспалительных реакций запускаются через рецепторы опознавания паттернов (pattern recognition receptors — PRR), которые содержатся как в клетках иммунной системы, так и в местном эпителии [25], и при связывании с микробными или эндогенными лигандами активируют внутриклеточные сигнальные пути синтеза провоспалительных цитокинов [25]. Тем не менее, результаты различных авторов, изучавших значение этих рецепторов и их каскадов в развитии индуцируемого представителями кишечной микрофлоры колоректального рака, противоречивы: одни свидетельствуют о протективной роли сигнальных путей Toll-подобных рецепторов (TLR) [28, 29], другие говорят об их проканцерогенном действии [30]. Вместе с тем мыши, лишенные гена NOD1/CARD4, кодирующего одноименный NOD-подобный рецептор, были более подвержены развитию колоректальных опухолей при воздействии декстрансульфата натрия в сравнении с обыкновенными мышами в аналогичных условиях [31], однако частота возникновения новообразований снижалась при антибиотикотерапии, пагубно влияющей на кишечную микрофлору. Запуск сигнальных каскадов, активируемых другими NOD-подобными рецепторами (NLRP3 и NLRP6), препятствует возникновению колоректального рака, поскольку у мышей, лишенных кодирующие эти рецепторы одноименных генов, злокачественные опухоли толстого кишечника развиваются чаще [32, 33].
По-видимому, усиленная активация рецепторов опознавания паттернов, возникающая при существенном изменении качественного и/или количественного состава микрофлоры, способна ингибировать канцерогенез, в то время как ухудшенное функционирование этой группы рецепторов может позволить трансформированным клеткам избежать опознавания иммунной системой и последующей гибели [25]. В то же время активация рецепторов опознавания паттернов может и промотировать канцерогенез в зависимости от внешних условий или особенностей самих рецепторов, создавая провоспалительное микроокружение, способствующее росту опухоли и развитию химиорезистентности и/или приводящее к иммуносупрессии, вызванной хроническим воспалением [25]. В этом случае, напротив, именно сниженная активность данных рецепторов минимизирует эффекты хронического воспаления и вероятность развития опухоли [25]. Таким образом, идеальным вариантом представляется «золотая середина» – соблюдение баланса между низкой и высокой активностью сигнальных путей, запускаемых с рецепторов опознавания паттернов [25]. Очевидно, что между путями вызываемого кишечной микрофлорой воспаления в толстом кишечнике существуют сложные взаимодействия, требующие дальнейшего детального изучения.
Вместе с тем роль иммунных механизмов в возникновении колоректального рака не вызывает сомнений. К примеру, J.M. Uronis и соавт. [34] показали, что у мышей, лишенных гена IL-10 (данный интерлейкин угнетает развитие воспаления) и экспонированных канцерогеном азоксиметаном, развиваются спонтанный колит и колоректальные карциномы, в то время как у обычных мышей, подвергшихся воздействию данного канцерогена, подобных изменений не наблюдается. Интересно, что у мышей-гнотобионтов, также лишенных гена IL-10 , после экспозиции азоксиметаном ни колита, ни опухолей также выявлено не было, что свидетельствует о ключевой роли представителей кишечной микрофлоры в развитии этих патологий [34]. Кроме того, у мышей-гнотобионтов, колонизированных исключительно Bifidobacterium animalis и лишенных гена IL-10 , в отличие от обычных мышей-гнотобионтов с подобной бактерией спустя некоторое время развивался колит умеренной степени тяжести [35]. Наконец, известно, что хронические воспалительные заболевания кишечника, такие, как язвенный колит и болезнь Крона, повышают риск развития колоректального рака [36]. Поскольку особенности микрофлоры кишечника существенно модулируют риск развития хронических воспалительных заболеваний кишечника [25], они также должны влиять на вероятность возникновения злокачественных опухолей ободочной и прямой кишки.
3. Механизмы, ассоциированные с канцерогенным действием бактериальных токсинов
Помимо метаболических нарушений, способствующих канцерогенезу, и модулирования иммунного ответа многие бактерии продуцируют токсины, обладающие канцерогенным действием через влияние на межклеточные взаимодействия, внутриклеточную передачу сигнала или индуцирование мутаций и эпигенетических изменений [37]. Таким образом, бактериальные токсины могут функционировать и как инициирующие, и как промотирующие агенты [37].
Известно, что энтеротоксигенные штаммы Bacteroides fragilis продуцируют фрагилизин [38], по своей сути являющийся металлопротеиназой, и рассекающий Е-кадгерин, который играет ключевую роль в регуляции контакта «клетка–клетка» и связан с внутриклеточными α- и β-катенинами. Деградация Е-кадгерина фрагилизином освобождает β-катенин, который мигрирует к ядру, где он активирует транскрипционные факторы, индуцирующие синтез белков c-myc и циклина D, обладающих разнонаправленным онкогенным действием [38]. Кроме того, этот токсин инициирует каскад активации транскрипционных факторов NF-κB и Stat3, что ведет к стимуляции синтеза провоспалительных цитокинов, промотирующих развитие опухоли [38]. Способностью вырабатывать обладающие канцерогенным действием токсины обладают и некоторые штаммы Escherichia coli, поскольку синтезируемый ими колибактин вызывает двуспиральные разрывы в ДНК, итогом чего могут являться онкогенные мутации [39]. Цитолетальный растягивающий токсин (cytolethal distending toxin – CDT), продуцируемый как минимум шестью видами грамотрицательных бактерий (в том числе и E. сoli), обладает аналогичным действием [40], а также приводит к повышению синтеза клетками слизистой оболочки кишечника многих провоспалительных молекул (TNF-α, NF-κB, IL-6, COX-2) и вызывает развитие дисплазии в модели микробного гепатоканцерогенеза [41]. Схожий механизм действия наблюдается у цикл-ингибирующего фактора (cycle inhibiting factor – Cif) многих патогенных штаммов E. coli, который дополнительно угнетает переход из фазы G2 в M-фазу жизненного цикла клетки, не влияя при этом на ДНК [42]. Помимо этого, другие штаммы E. coli продуцируют цитотоксический некротизирующий фактор 1 (cytotoxic necrotizing factor 1 – CNF1), активирующий белки семейства Rho, которые характеризуются ГТФ-азной активностью и являются ключевыми для регуляции многих витальных процессов (пролиферации, выживания, апоптоза, ангиогенеза, подвижности и других) [43]. Это, в свою очередь, запускает ряд внутриклеточных сигнальных путей, включая те, которые функционируют через потенциально важные для неопластической трансформации белки FAK, Src, COX-2 и NF-κB, итогом чего является ускоренное деление клетки, ее резистентность к апоптозу, усиленный синтез провоспалительных цитокинов и другие онкогенные нарушения [44]. Наконец, липополисахарид, являющийся основным гликолипидным компонентом наружной мембраны грамотрицательных бактерий, способствует адгезии и инвазии опухолевых клеток через связывание с Тоll-подобным рецептором 4 (TLR4), что, в свою очередь, ведет к активации NF-κB и повышению синтеза урокиназного плазминогенного активатора (uPA) [45]. Данный белок катализирует преобразование плазминогена в плазмин, деградирующий белки межклеточного матрикса, в результате чего миграция и адгезия злокачественных клеток ускоряется [45].
4. Связь отдельных представителей кишечной микрофлоры с риском развития колоректального рака
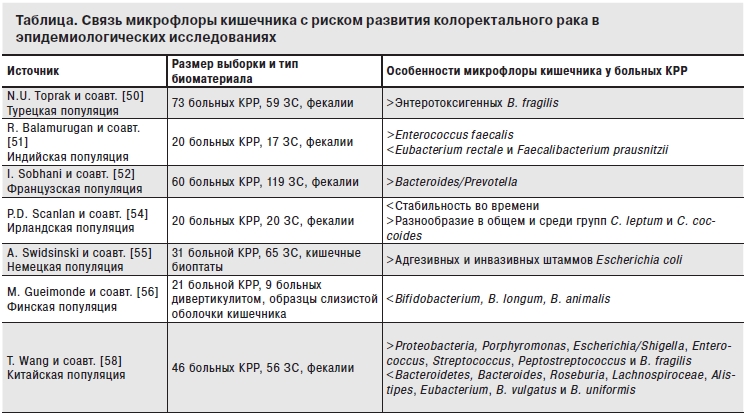
В последние годы количество экспериментальных и особенно эпидемиологических работ, посвященных данному вопросу, значительно возросло. В одном из первых проведенных по этой проблеме исследований W.E. Moore и соавт. [46] обнаружили в фекалиях больных полипозом повышенное содержание Bacteroidesvulgatus, Eubacteriumspp., Ruminococcusspp., Streptococcushansenii, Bifidobacteriumspp. и Faecalibacteriumprausnitzii, а также сниженную концентрацию LactobacillusS06 и Eubacteriumaerofaciens. В нескольких исследованиях [34, 47, 48] было доказано, что у мышей-гнотобионтов риск развития колоректального рака при врожденных генетических нарушениях или при экспозиции канцерогенами значительно ниже, чем у мышей, колонизированных нормальной микрофлорой. Так, S. Wu и соавт. [49] показали, что у мышей, искусственно колонизированных энтеротоксигенными штаммами Bacteroides fragilis на протяжении длительного времени, развивается вначале колит, а затем злокачественные опухоли толстого кишечника. Полученные на этой животной модели результаты были далее подтверждены N.U. Toprak и cоавт. [50], выявившими энтеротоксигенные штаммы данной бактерии в фекалиях 38% больных колоректальным раком (в контрольной группе этот показатель составил 12%, ОР=4,16, 95%ДИ=1,39–12,43, p=0,009). R. Balamurugan и соавт. [51] обнаружили, что у больных колоректальным раком снижено содержание в кишечнике таких бактерий, как Eubacterium rectale и Faecalibacterium prausnitzii, и повышена концентрация Enterococcus faecalis в сравнении с контрольной группой. По данным I. Sobhani и соавт. [52], у пациентов со злокачественными опухолями ободочно и прямой кишки также повышена концентрация представителей родов Bacteroides и Prevotella. A. Boleij и соавт. [53] выявили, что у пациентов с полипами и I–II стадиями колоректального рака наблюдается более сильный иммунный ответ на антиген Streptococcus bovis RpL7/L12 в сравнении с асимптоматичными субъектами. P.D. Scanlan соавт. [54] обнаружили, что микрофлора кишечника больных колоректальным раком и полипозом характеризуется сниженной стабильностью во времени и повышенным разнообразием в сравнении с кишечной микрофлорой в контрольной группе. Кроме того, в двух опытных группах также было отмечено повышенное разнообразие внутри видов Clostridium leptum и Clostridium coccoides [54]. В исследовании, проведенном A. Swidsinski и соавт. [55], в слизистой толстого кишечника больных колоректальным раком в 82% случаев были идентифицированы адгезивные и инвазивные штаммы Escherichia coli, в то время как в контрольной группе этот показатель был статистически достоверно ниже. Аналогичная картина была обнаружена при сравнении тканей колоректальных аденом и слизистой оболочки толстого кишечника в контрольной группе [55]. M. Gueimonde и соавт. [56] выявили, что у больных колоректальным раком и воспалительными заболеваниями кишечника наблюдается меньшее содержание Bifidobacterium spp., а также видов Bifidobacteriumlongumи Bifidobacteriumanimalis по сравнению с пациентами с дивертикулитом. По мнению авторов, это может свидетельствовать о том, что патологические изменения микрофлоры могут быть выявлены на самых ранних стадиях цепи многоступенчатого канцерогенеза [56]. X.J. Shen и соавт. [57] показали, что в слизистой оболочке кишечника пациентов с колоректальными аденомами содержание микроорганизмов, относящихся к типу Proteobacteria, повышено, а относящихся к типу Bacteroidetes – снижено. При анализе более мелких таксономических групп было обнаружено, что бактерии Dorea spp., Shigella spp. и Faecalibacterium spp. встречаются в тканях аденом чаще, а Bacteroidesspp. и Coprococcusspp. – реже в сравнении с аналогичными нормальными тканями субъектов контрольной группы [57]. Кроме того, количество и разнообразие бактерий в аденомах было выше, чем в соответствующих тканях субъектов контрольной группы [57]. Результаты, полученные Т. Wang и соавт. при сравнении бактериального состава фекалий больных колоректальным раком и пациентов контрольной группы [58], подтвердили данные X.J. Shen et al. в отношении типов Proteobacteria и Bacteroidetes. В этом исследовании у пациентов со злокачественными опухолями ободочной и прямой кишки было повышено количество бактерий родов Porphyromonas, Escherichia/Shigella, Klebsiella, Enterococcus, Streptococcus и Peptostreptococcus и снижено содержание микроорганизмов Bacteroides, Roseburia, Lachnospiroceae, Alistipes, Eubacteriumи Parasutterella[58]. Интересно, что в отношении распространенности различных видов бактероидов в изучаемых группах наблюдались существенные различия: так, концентрация Bacteroides fragilis была статистически достоверно выше в фекалиях больных колоректальным раком, а содержание Bacteroides vulgatus и Bacteroides uniformis – в фекалиях субъектов контрольной группы [58]. J.R. Marchesi и соавт. [59] показали, что в образцах злокачественных колоректальных новообразований была повышена концентрация бактерий Coriobacteridae, Roseburia, Fusobacterium и Faecalibacterium и снижено содержание представителей семейства Enterobacteriaceae (Citrobacter spp., Shigella spp., Cronobacter spp., Kluyvera spp., Serratia spp. и Salmonella spp.) в сравнении с окружающими здоровыми тканями. Авторы предположили, что поскольку указанные первыми группы бактерий обладают способностью продуцировать короткоцепочечные жирные кислоты, характеризуются пробиотическими свойствами и являются комменсалами, их повышенная концентрация в опухолевых тканях объясняется тем, что они колонизируют их уже после возникновения заболевания, вытесняя бактерии второй группы, которые в большинстве своем обладают некоторыми канцерогенными свойствами [59]. Таким образом, теоретически возможно участие микрофлоры в предотвращении распространения опухоли уже после ее появления [59]. В то же время привлечение бутират-продуцирующих бактерий в опухолевый очаг может, напротив, способствовать развитию опухоли, т. к. бутират, обладая антиканцерогенным потенциалом, одновременно является и источником энергии для опухолевых клеток, а также ослабляет иммунный ответ, позволяя опухолевым клеткам избегать уничтожения [59]. Наконец, в одной из последних работ M. Castellarin и соавт. [60] продемонстрировали, что в тканях злокачественных опухолей ободочной и прямой кишки содержание Fusobacterium nucleatum значительно выше (в 79–415 раз), чем в соответствующих нормальных тканях, причем у субъектов с более высокой концентрацией этого вида бактерий в опухолевых тканях был повышен риск метастазирования в регионарные лимфатические узлы. Краткие характеристики указанных выше исследований по связи представителей микрофлоры кишечника с риском развития колоректального рака приведены в таблице.
Примечание. КРР – колоректальный рак, ЗС – здоровые субъекты.
Выводы
Исходя из описанных выше механизмов канцерогенеза очевидно, что особенности качественного и количественного состава микрофлоры толстого кишечника могут оказывать существенное влияние на вероятность возникновения колоректального рака. Проведенные в последние годы экспериментальные и популяционные исследования позволили предположить и обосновать связь ряда ее представителей с риском развития этого заболевания. В то же время проведенных работ явно недостаточно для того, чтобы говорить о подробном изучении данной проблемы. Помимо новых клеточных и животных моделей биологического канцерогенеза необходимы и широкомасштабные эпидемиологические исследования в различных странах для сравнения межпопуляционных различий влияния микрофлоры на риск развития злокачественных опухолей ободочной и прямой кишки. Важно отметить, что биологическими материалами для анализа должны быть не только фекалии, но и образцы слизистой оболочки кишечника, поскольку состав микрофлоры в этих двух типах биоматериалов различается. Кроме того, зачастую вследствие недостаточно тщательно разработанного дизайна исследования бывает сложно сделать вывод о том, играет ли изучаемый представитель микрофлоры толстого кишечника роль в этиологии колоректального рака или всего лишь колонизирует опухоль после ее возникновения в результате нарушений местного гомеостаза. Во избежание подобной проблемы следует включать в исследование не только больных злокачественными опухолями ободочной и прямой кишки и контрольную группу из здоровых людей, но и как минимум две дополнительные контрольные группы: пациентов с хроническими воспалительными заболеваниями кишечника (язвенный колит, болезнь Крона) и пациентов с полипозом толстой кишки, поскольку эти патологии являются предраковыми состояниями по отношению к колоректальным злокачественным новообразованиям. В идеале все субъекты опытной и трех контрольных групп должны быть сопоставимы по полу, возрасту и региону постоянного проживания, однако из-за технических сложностей представляется возможным допустить и лишь приблизительное сходство обследуемых по возрасту (±2–3 года). Сравнение особенностей микрофлоры толстого кишечника у обозначенных четырех групп позволяет в относительно полной мере проследить их изменения по стадиям многоступенчатого канцерогенеза, что имеет важное значение как для определения статуса представителя микрофлоры (является он этиологическим агентом злокачественных опухолей или нет), так и для профилактики, поскольку в этом случае по изменениям качественного и количественного состава микрофлоры можно сделать вывод о вероятности развития колоректального рака и, следовательно, выявить группы риска.
Отдельно стоит отметить множественные взаимодействия между типом питания, микрофлорой толстого кишечника и риском возникновения колоректальных злокачественных новообразований, поскольку состав употребляемых в пищу продуктов напрямую влияет на состав микрофлоры, что, соответственно, модулирует вероятность развития рака. Изучение таких взаимодействий для определения оптимального для профилактики колоректального рака типа питания, несомненно, является перспективным направлением по отношению к рассматриваемой проблеме. Более того, по-видимому, место типа питания в такой триаде могут занимать и другие факторы риска развития рака: табакокурение, экспозиция профессиональными канцерогенами и канцерогенами окружающей среды, низкая физическая активность, нарушения гормонального фона, прием некоторых химиопрепаратов и т.д. С другой стороны, микрофлора толстого кишечника может и сама влиять на другие факторы риска развития рака, к примеру, на ожирение, образование канцерогенных метаболитов, развитие местного и системного воспаления или на синтез онкогенных белков.
Наконец, еще одним актуальным как с теоретической, так и с практической точки зрения вопросом является возможная роль пребиотиков, пробиотиков и синбиотиков в профилактике злокачественных опухолей толстой и прямой кишки, в значительной степени вызванных биологическими агентами. Для его решения оптимально сочетание результатов экспериментальных работ на животных моделях биологического канцерогенеза и эпидемиологических проспективных исследований, которые, однако, требуют длительного времени для осуществления. Такой способ профилактики может оказаться весьма эффективным и привести к существенному снижению экономических потерь, заболеваемости, инвалидизации и смертности от колоректального рака.
Новые выявленные в будущем корреляции между представителями кишечной микрофлоры и риском развития колоректального рака могут быть использованы в интегративных системах определения риска развития злокачественных новообразований ободочной и прямой кишки на основе этих связей, которые, в свою очередь, целесообразно будет использовать в различных профилактических программах. Более того, поскольку многие механизмы ассоциированного с микрофлорой толстого кишечника биологического канцерогенеза являются системными, возможно предположить роль микрофлоры толстого кишечника в возникновении и других (экстраинтестинальных) типов злокачественных опухолей. Аналогичная гипотеза должна быть справедлива и для микрофлоры других органов в случае выявления у нее канцерогенных механизмов системного действия. Актуальность данной проблемы подчеркивается ее малоизученностью и несомненной перспективностью с теоретической и практической позиций. Судя по наметившейся в последние годы тенденции к увеличению доли случаев рака, ассоциированных с биологическими канцерогенами, а также благодаря непрерывному прогрессу методов молекулярной биологии в будущем, вероятно, будет открыто множество связей представителей микрофлоры тела человека с риском развития различных типов злокачественных новообразований.

{kind=link}