Одним из биотических факторов среды, которые могут оказывать влияние на напряженность очагов крымской-конго геморрагической лихорадки (ККГЛ), является присутствие в организме переносчика дополнительного агента вирусной этиологии. Это обстоятельство может привести к изменению обычных симбиотических взаимоотношений между вирусом ККГЛ и его основным переносчиком — клещом Hyalomma marginatum marginatum [1]. На территории Астраханской области существуют сочетанные очаги чумы, туляремии, риккетсиозов и арбовирусных инфекций [2]. Наличие единых
переносчиков и механизмов передачи создает условия для совместной циркуляции возбудителей, при этом увеличивается риск возникновения микстинфекций. По данным многолетних исследованиий, проведенных в Астраханской области, была установлена циркуляция вируса Баткен/Дхори (Б/Д) — представителя рода Thogotovirus семейства Orthomyxoviridae [13]. Показатель зараженности клещей Hyalomma marginatum marginatum вирусом Б/Д в этой области в отдельные годы был значительно выше, чем вирусом ККГЛ [1]. В 1965—1966 гг. из клещей Hyalomma plumbeum plumbeum (их современное
название — Hyalomma marginatum marginatum), собранных в Астраханской области, был выделен вирус Астра, имевший родство с вирусом Дхори [4, 5]. В последующие годы при изучении циркуляции арбовирусов на территории этой области были отмечены находки вируса Дхори [6].
В настоящем сообщении приведены данные о распространении вируса Б/Д на территории
Астраханской области РФ и возможном его влиянии на спонтанную инфицированность клещей
Hyalomma marginatum marginatum вирусом ККГЛ (по материалам 1980—1990 и 2005—2009 гг.).
Материалы и методы
Иксодовых клещей собирали с домашних животных в стационарных пунктах 10 районов Астраханской области подекадно в период массового хода членистоногих (апрель—май). Всего исследовано 43 724 клеща, из которых было приготовлено 4335 суспензий. Формирование проб, обработку клещей для вирусологических исследований проводили, как описано ранее [7]. Для определения антител к вирусу Б/Д были исследованы пробы сыворотки крови 548 сельских жителей области и 1328 коров.
При исследовании вирусофорности клещей использовали метод внутримозгового заражения новорожденных белых мышей 2-суточного возраста (НБМ). Результаты заражения оценивали
с помощью иммуноферментного анализа (ИФА) (определение антигена вируса ККГЛ в суспензии
мозга НБМ) и реакции связывания комплемента (РСК) (определение антигенов вируса ККГЛ
и вируса Б/Д) [7, 8]. Для 7 штаммов Б/Д и 16 штаммов ККГЛ, изолированных в 2005—2009 гг.,
их идентификация была подтверждена с помощью специфической полимеразной цепной реакции
(ПЦР) [9].
Для изучения патогенности вируса Б/Д использовали лабораторных животных: НБМ, молодых белых мышей массой 5—6 г (МБМ), взрослых белых мышей массой 18—20 г (ВБМ), морских свинок массой 300—400 г. Для изучения взаимодействия вируса Б/Д с клетками млекопитающих использовали культуру клеток Vero-E6. Для серологических исследований готовили сахарозоацетоновые антигены, используя штаммы К251-243 (Астраханская область, 1990) и Ходжа (Узбекистан, 1967) вируса ККГЛ и штаммы К251-55 (1990 г.) и К379-106 (2005 г.) вируса Б/Д, выделенные в Астраханской области.
Применяли метод внутримозгового и внутрибрюшинного заражения НБМ, МБМ, ВБМ и внутрибрюшинный метод заражения морских свинок. Заражение культуры клеток и выявление колоний бляшек под агаровым покрытием [10], обработку эфиром и дезоксихолатом натрия [11], постановку
реакции гемагглютинации [3] и РСК [8] выполняли, как описано ранее.
Геном штаммов вируса Б/Д был охарактеризован путем прямого секвенирования нуклеотидных последовательностей фрагментов гена гликопротеина с праймерами DHV-G1 CACTTTGCCTACCACACGTGCA ACCTAT и DHV-G2 CCGAGCAATGTTCCCAACAGTCTGT (577 нуклеотидов, сегмент 4) и гена нуклеопротеина с праймерами DHV-N1 CTTGTGGCAGCGGAACCCAGTCA и DHV-N2
GCAGGCAGCGAAAGGTGCAGTAGATTA (486 нт, сегмент 5). Праймеры DHV-G1 и DHV-G2 использовали также в ПЦР для идентификации вируса Б/Д.
Результаты и обсуждение
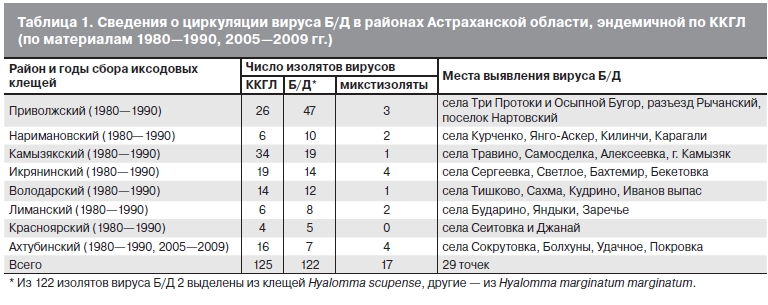
Данные о циркуляции вируса Б/Д, полученные нами в сезоны 1980—1990 гг., свидетельствовали о широком распространении этого арбовируса в дельтовых (южных) районах Астраханской области. Вирус Б/Д был выявлен в 29 точках сбора иксодовых клещей на территории 7 районов, расположенных в дельте Волги. В годы высокой численности иксодовых клещей (1983—1985, 1989) было отмечено значительное увеличение числа суспензий, содержавших вирус Б/Д. Основным источником выделения вируса Б/Д были клещи H. marginatum marginatum. Чаще находки этого вируса имели место на территории Приволжского, Камызякского, Икрянинского, Володарского и Наримановского районов (от 10 до 47 изолятов). Меньшее число таких находок (5—8 изолятов) приходилось на Лиманский, Красноярский районы и 1 изолят был получен в этот период из материалов, собранных в Ахтубинском районе. Исследования, проведенные в 2005—2009 гг. в северном Ахтубинском районе, свидетельство-
вали об активной циркуляции вируса Б/Д и на данной территории (табл. 1).
Таблица 1. Сведения о циркуляции вируса Б/Д в районах Астраханской области, эндемичной по ККГЛ
(по материалам 1980—1990, 2005—2009 гг.).
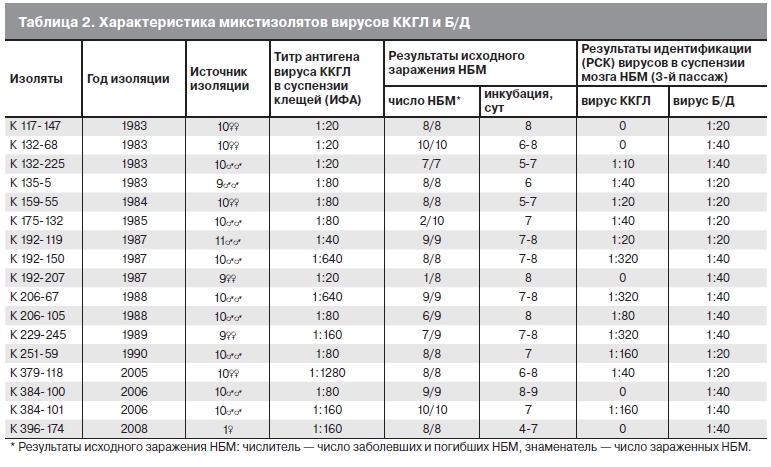
На протяжении многих лет наблюдения за очагом ККГЛ при исследовании суспензий иксодовых клещей регулярно выявляли микстизоляты, содержавшие по крайней мере 2 вируса: ККГЛ и Б/Д. В табл.2 представлены результаты исследования 17 микстштаммов, выделенных из клещей H. marginatum marginatum. Основная часть этих пулов клещей (11 проб) состояла из самцов (9—10 экземпляров в пробе), другая часть (6 проб) — из самок, при этом 5 проб содержали по 9—10 клещей и 1 проба — 1 клеща. Штамм К396-174, выделенный из одной питающейся самки массой 0,7 г, является достоверным свидетельством присутствия двух вирусов в одном клеще, отловленном в природе. Все исходные
суспензии иксодовых клещей содержали антиген вируса ККГЛ, выявляемый с помощью ИФА. При
первичном заражении НБМ суспензиями клещей в 12 случаях наблюдали 100% заболевших НБМ
спустя 5—8 сут после внутримозгового заражения, в 5 случаях больные мыши составили 20—70%
от числа инфицированных. При идентификации выделенных агентов в суспензиях мозга больных
НБМ с помощью РСК были определены антигены вирусов ККГЛ и Б/Д. Микстизоляты, выделенные в сезоны 1980—1990 гг., были пассированы не более 3 раз (срок наблюдений) через мозг НБМ с подтверждением в РСК присутствия двух вирусов. Результаты идентификация вирусов Б/Д и ККГЛ в суспензиях мозга больных НБМ на уровне 3-го пассажа свидетельствовали о наличии двух вирусов в 12 из 17 исследованных проб (см. табл. 2). В суспензиях мозга НБМ, зараженных 5 микстштаммами, на уровне 3-го пассажа определяли антиген только вируса Б/Д. В 5 случаях титры антигена вируса ККГЛ были в 4—8 раз выше, чем вируса Б/Д. В 7 других случаях титры антигенов обоих вирусов составляли 1:10—1:80.
Таблица 2. Характеристика микстизолятов вирусов ККГЛ и Б/Д.
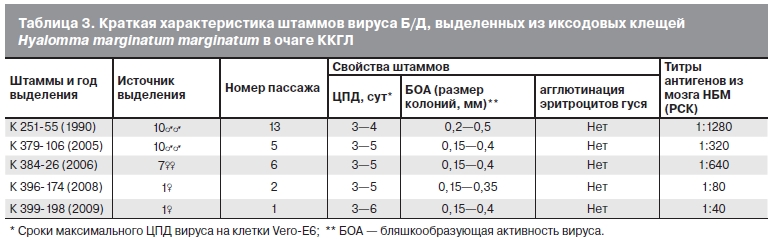
Краткая характеристика свойств вируса Б/Д представлена в табл. 3 на примере отдельных штаммов,
выделенных в разные годы. Все изученные нами штаммы вируса Б/Д вызывали летальную инфекцию у НБМ, МБМ и ВБМ после внутримозгового и внутрибрюшинного заражения. Инкубационный период после первичного заражения составлял 7—8 сут, через 3—5 пассажей через мозг НБМ он составил 4—5 сут. Титры вируса в суспензиях мозга зараженных НБМ на уровне 3—5-го пассажей достигали 6,5 lg ЛД /0,02 мл, 5,5 lg ТЦД /0,1 мл и 6,0 lg БОЕ /0,1 мл. Штаммы вируса Б/Д были чувствительны к действию жирорастворителей, не вызывали агглютинацию эритроцитов гуся при значениях рН в диапазоне от 5,2 до 7,6. В культуре клеток Vero-E6 отмечено размножение вируса Б/Д с развитием выраженного цитопатогенного действия (ЦПД), завершающегося полным разрушением клеток, под агаровым покрытием регистрировали колонии-бляшки, размеры которых в течение срока наблюдений увеличивались от 0,15 до 0,5 мм (см. табл. 3). Титры вируса Б/Д в культуральной жидкости зараженных культур клеток составляли 6,5—7,0 lg ТЦД /1 мл. Сахарозоацетоновые антигены, приготовленные из мозга зараженных НБМ на уровне 3—5-го пассажей, имели титры от 1:80 до 1:1280. В сыворотках морских свинок, получивших не менее 3 инъекций вируса, выявлялись комплементсвязывающие антитела к вирусу Б/Д в титре от 1:32 до 1:1024.
Таблица 3. Краткая характеристика штаммов вируса Б/Д, выделенных из иксодовых клещей.
Секвенирование двух штаммов Б/Д, изолированных в Астрахани, показало, что они наиболее генетически близки к прототипному штамму Баткен LEIV306K (гомология 95% по нуклеотидной последовательности и 99,4% по аминокислотной последовательности исследованных фрагментов гликопротеина и нуклеопротеина). От прототипного штамма 1313/61 вируса Дхори они отличаются на 17 и 6,5% по нуклеотидной и аминокислотной последовательности соответственно. При этом их отличие от штаммов вируса вида Тогото, принадлежащего тому же роду Thogotovirus, составляет 52—54% по нуклеотидной и 60—62% по аминокислотной последовательности (см. рисунок).
Рисунок. Филогенетическое дерево вирусов рода Thogotovirus.
Антитела к вирусу Б/Д в титре 1:8—1:32 были найдены у 4—6,5% сельских жителей дельтовых районов области и у 1,2—6,3% обследованных домашних животных (коров). Следует отметить, что в пробах
сыворотки крови коров (n=541) из Ахтубинского района, полученных в мае 2009 г., были определены антитела к вирусам ККГЛ (1,1%) и Б/Д (2%); у 0,3% обследованных животных были обнаружены антитела к двум указанным вирусам.
Широкое распространение природно-очаговых микстинфекций, передающихся клещами, обусловлено закономерностью взаимоотношений различных возбудителей в организме переносчика и в экосистеме в целом [12]. Результаты 13-летнего мониторинга за очагом ККГЛ на территории Астраханской области показали, что высокая численность иксодовых клещей H. marginatum marginatum, как правило, сочетавшаяся с высокими показателями вирусофорности членистоногих, является одним из биотических факторов, оказывающих влияние на состояние активности очага ККГЛ. Эпизоотии среди диких млекопитающих — прокормителей неполовозрелых клещей могут приводить к депрессии численности клещей, в том числе и вирусофорных особей. За время наблюдения был отмечен 5-летний период (1983—1987) умеренного снижения спонтанной инфицированности клещей H. marginatum marginatum вирусом ККГЛ, который совпал или был результатом резкого подъема уровня зараженности этих клещей вирусом Б/Д. Даже незначительные подъемы спонтанной инфицированности клещей вирусом
Б/Д совпадали со снижением уровня зараженности этих членистоногих вирусом ККГЛ [1]. Феномен
«ножниц» в динамике показателей инфицированности клещей H. marginatum marginatum вирусами
ККГЛ и Б/Д предполагает «соперничество» этих арбовирусов в организме переносчика. Широкое
распространение вируса Б/Д в очаге ККГЛ может оказаться важным фактором регулирования доли
H. marginatum marginatum, спонтанно зараженных вирусом ККГЛ.
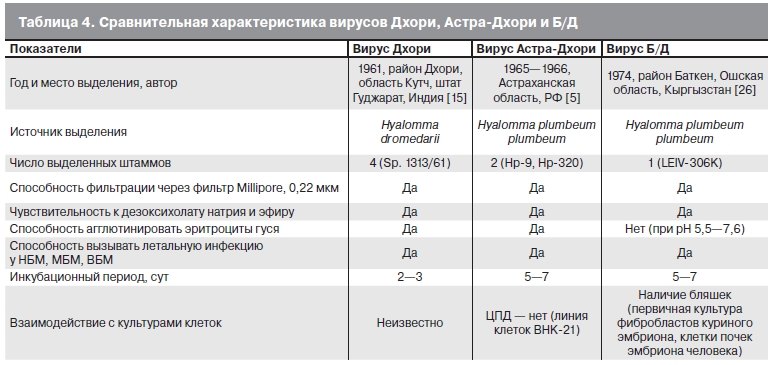
В связи с имеющимися данными о родстве вируса Б/Д с вирусом Дхори [13, 14] приводим краткую сравнительную характеристику этих агентов (табл. 4). Вирус Дхори впервые был выделен в Индии из клещей Hyalomma dromedarii, снятых с верблюдов в районе Дхори, область Кутч, штат Гуджарат. Антитела к вирусу Дхори были выявлены у местного населения, а также у лошадей [15]. Позже антитела к этому вирусу были определены в пробах сыворотки людей и домашних животных из 6 штатов и 1 союзной территории Индии [16]. Циркуляция вируса Дхори была зарегистрирована в Египте, Кении, Саудовской Аравии, Пакистане и Португалии [17—20]. Штаммы вируса Астра, выделенные в Астраханской области, Краснодарском крае, Азербайджане и Армении, агглютинировали эритроциты гуся и имели антигенные связи между собой и с вирусом Дхори [4, 5, 21—23]. В 2001—2007 гг. на юге России вирус Дхори был выделен от клещей H. marginatum marginatum, зайца-русака и большого баклана; антитела к этому вирусу обнаружены у жителей в дельте реки Кубань [6, 24, 25].
Прототипный штамм LEIV-306K вируса Баткен был выделен из имаго H. plumbeum plumbeum, снятых в апреле 1970 г. с овец в районе Баткен, Ошская область, Кыргызстан [26]. Этот вирус фильтровался
через поры 0,22 мкм, содержал РНК, был чувствителен к действию эфира и дезоксихолата натрия, не агглютинировал эритроциты гуся (см. табл. 4). Спустя 10 лет стало известно об изоляции вируса Баткен
из иксодовых клещей, собранных в Узбекистане [27]. В 1988 г. было сообщено о результатах идентификации вирусных агентов, выделенных из иксодовых клещей, собранных в 1980—1984 гг. при обследовании очагов ККГЛ на территориях Астраханской области и Крыма. В опытах заражения НБМ был изолирован 51 штамм: 50 — из клещей, собранных в Астраханской области, и 1 — из клещей, собранных в Крыму. Эти штаммы вызывали летальное заболевание у НБМ, МБМ и ВБМ, были чувствительны к действию жирорастворителей, не агглютинировали эритроциты гуся, в своей структуре содержали РНК. В культуре клеток 6619-1, зараженных вирусом Баткен, через 72—96 ч отмечали деструкцию клеток, заканчивающуюся полным их разрушением. По результатам РСК выделенные штаммы имели между собой перекрестные антигенные связи и давали четкую положительную реакцию с антисывороткой к штамму LEIV-306K вируса Баткен [3]. Позднее на основании данных серологических исследований, электронной микроскопии [13] и результатов молекулярно-биологического анализа [14] было установлено, что штамм LEIV-306K имеет родство с вирусом Дхори; согласно международной классификации (www.ncbi.nlm.nih.gov/taxonomy, www.ictvonline.org/virusTaxonomy.asp?version=2009), они относятся к одному виду. Действительно, генетическое различие в группе штаммов Дхори, Баткен и Б/Д составляет порядка 10,4±0,6%, т. е. соответствует внутривидовому, в то время как их отличие от группы штаммов вида Тогота превышает 50% (см. рисунок). Однако результаты сравнительной характеристики свойств вирусов Дхори, Астра- Дхори и Б/Д, представленные в табл. 4, и выделение вирусов Б/Д в отдельную ветвь на филогенетическом дереве свидетельствуют о некоторых различиях между вирусами Дхори (и Астра-Дхори), с одной стороны, и вирусом Б/Д, с другой стороны. Имеются также данные об отсутствии гемагглютинирующей активности изолятов вируса Баткен, полученные разными исследователями в Крыму и Астраханской области РФ [3, 28], Кыргызстане [29], Азербайджане [13]. Приведенные выше сведения позволяют рассматривать группу щтаммов Б/Д как подтип вида Дхори, отличный от штамма 1313/61.
Таблица 4. Сравнительная характеристика вирусов Дхори, Астра-Дхори и Б/Д.
Роль вирусов Дхори, Б/Д и Астра-Дхори в инфекционной патологии человека и животных пока
неясна. Описаны клинические случаи заболевания, возникшие в результате лабораторного заражения вирусом Дхори, штамм 1313/61, вероятно, аэрогенным путем. Заболевание сопровождалось высокой лихорадкой, головными и мышечными болями, симптомами поражения ЦНС, лейкопенией. Клинические симптомы болезни регрессировали в течение 2—3 нед без каких-либо остаточных явлений, но у всех больных длительно (до 1—2 мес) сохранялся выраженный астенический синдром [30]. Следует отметить, что летальная инфекция, развивавшаяся после интраназального заражения взрослых мышей вирусом Дхори, также сопровождалась лейкопенией и тромбоцитопенией, резким повышением концентрации фактора некроза опухоли, интерлейкинов-1, 6, 10 и других провоспалительных цитокинов и хемокинов, поражением легких, печени, мозга [31]. Известно, что вирус Тогото, также переносимый иксодовыми клещами, способен вызвать лихорадку и/или менингоэнцефалит у человека [32—34].
Изучение очага ККГЛ, проведенное нами в Астраханской области в сезоны 1980—1990 и 2005—2009 гг., выявило интенсивную циркуляцию вируса Б/Д, что является благоприятным условием для появления микстизолятов. Единый переносчик и его прокормители создают условия для совместной циркуляции этих двух вирусов как в разных особях клещей, так и в одном членистоногом. В наших исследованиях изолят К 396-174, полученный из одной самки H. marginatum marginatum, содержал вирусы Б/Д и ККГЛ, что позволяет достоверно утверждать наличие микстинфекции. Другие 16 изолятов были выделены
из суспензий клещей этого же вида, содержавших по 9—10 экземпляров членистоногих. Не исключено, что вирусами Б/Д и ККГЛ были инфицированы разные особи клещей. В результате последующих
пассажей через мозг НБМ такие изоляты стали микстизолятами, содержащими 2 вируса — ККГЛ
и Б/Д, что подтверждено данными ИФА, РСК и ПЦР. Полученные нами данные позволяют предполагать в некоторых случаях невыгодные для вируса ККГЛ условия размножения в присутствии вируса Б/Д.
Таким образом, широкое распространение вируса Б/Д может оказывать двойственное влияние на инфекционную заболеваемость человека. С одной стороны, вирус сам потенциально способен вызывать заболевания, и это значит, что в дальнейшем адекватные методы, способные выявлять эту инфекцию, должны применяться для диагностики лихорадок и энцефалитов неясной этиологии на юге России и в Средней Азии. С другой стороны, не исключено, что присутствие вируса Б/Д в клещах и их прокормителях отчасти ограничивает циркуляцию более опасного вируса ККГЛ, что должно быть изучено в первую очередь в экспериментальных условиях.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}