
Основными возбудителями острых гнойных менингитов (ОГМ) являются Neisseria meningitidis, Streptococcus pneumoniae, Haemophilus influenzae,Escherichia coli, Streptococcus agalactiae (серогруппы В). Эти микроорганизмы принадлежат к далеким друг от друга родам и даже семействам, но, вместе с тем, их сближают некоторые общие свойства. Так, они живут в природе как внеклеточные микроорганизмы, т. е. формируют сообщества на поверхности клеток макроорганизма. Второе, не менее удивительное для возбудителей ОГМ обстоятельство – они представлены преимущественно не как паразиты, а как безвредные для человека комменсалы, заселяющие слизистые оболочки вместе с другими представителями нормальной флоры. Даже такой безусловный патоген, как N. meningitidis может десятилетиями циркулировать среди населения в форме бессимптомной колонизации носоглотки (здоровое носительство). Первые 3 из вышеназванных микроорганизмов обитают в верхних дыхательных путях человека, 2 последних – в кишечнике. Третье свойство, общее для этих «безвредных комменсалов» – капсулообразование, важнейшая характеристика патогенности, присущая далеко не всем классическим возбудителям. Капсула защищает бактериальных паразитов от их главных врагов – фагоцитов, антител и лизирующего действия комплемента. Однако капсулообразование может реализоваться микроорганизмом преимущественно при попадании его во внутреннюю среду организма. А попасть туда непросто, так как неповрежденные естественные и иммунологические барьеры человека непроницаемы для комменсалов. В этом сообщении мы намерены кратко осветить процесс проникновения основных возбудителей ОГМ в организм хозяина вплоть до субарахноидального пространства головного мозга, обращая внимание на недавно выявленные особенности патогенеза.
Первым этапом атаки инфекционного агента является колонизация слизистой оболочки в месте входных ворот, осуществляемая с помощью прочного прикрепления (адгезии) микроорганизма к эпителию. Этому предшествует противостояние движению ресничек, кашлевым актам, потокам слизи, а также антагонистическому действию нормальной микрофлоры. Однако в ряде случаев закрепившийся на слизистой оболочке микроорганизм проникает в подслизистый слой, вызывая местный воспалительный процесс, еще реже – в кровь, а оттуда на мозговые оболочки (рис. 1, см. на вклейке). Важную роль в инвазии бактерий с поверхности слизистых оболочек играют факторы врожденного и приобретенного иммунитета человека [1].
Процесс колонизации и последующей инвазии в кровь, вплоть до мозговых оболочек, наиболее полно изучен у N. meningitidis. К концу прошлого века стал ясен механизм начальных взаимодействий инфекционных агентов (в том числе возбудителей ОГМ) с организмом хозяина [2–4]. Этот возбудитель попадает с аэрозолем на слизистую оболочку носоглотки и колонизирует ее, т. е. создает плацдарм для обеспечения своего дальнейшего распространения. Менингококк, попавший от источника на слизистую оболочку нового лица, часто имеет капсулы, защищающие его от высыхания и других факторов внешней среды. Однако капсулы мешают бактериальным клеткам прикрепляться к эпителию и друг к другу, так как экранируют (заслоняют) собой липкие поверхностные структуры – адгезины. Поэтому в носоглотке приживляются штаммы, не экспрессирующие капсул, или генетически лишенные этой способности. Бактериологи знают, что из носоглотки обычно высеваются бескапсульные формы менингококка. Прикрепление вначале идет за счет пилей. Пили – отходящие от оболочки бактерии трубочки шириной около 6 нм и длиной несколько мкм, состоящие из белков пилинов (Pil) с разными свойствами. Основной ствол состоит из белка пилина PilE. Пили не только прикрепляют клетки менингококка к эпителию (за счет концевого липкого пилина PilC) и друг к другу, но и способствуют расползанию возбудителя по слизистой оболочке (за счет пилина PilX [5]). Более того, по трубочкам в клетку менингококка поступают фрагменты ДНК других распавшихся бактерий своего, а иногда и иного вида (одна из причин генетической изменчивости менингококка – трансформация). В сцеплении клеток друг с другом участвует белок пилей PilU [6].
Рецепторы на эпителии для пилинов до сих пор неизвестны. Ранее эту функцию приписывали мембранному белку CD46, с чем сейчас согласны далеко не все [3]. Дальнейшее взаимодействие возбудителя и человека, в зависимости от штамма и уровня иммунной защиты, может развиваться двояким путем. Чаще всего на эпителии образуются микроколонии, которые сливаются и распространяются вширь благодаря «дергающейся» подвижности менингококковых клеток, обеспечивающейся сократительными движениями пилей. При этом находящиеся на поверхности слизистой оболочки менингококки иногда проникают вглубь эпителия и снова выходят наружу [7]. Менингококк выделяет во внешнюю среду полисахариды, белки, липиды, продукты жизнедеятельности, образующие плотную структуру – матрикс, в которую погружены планктонные бактерии. Таким путем на слизистой оболочке образуется многослойная плотная биопленка [8]. Она прочно прикреплена к поверхности эпителия с помощью липкой экзогенной ДНК, освобождающейся при аутолизе части бактериальных клеток менингококка [9]. В биопленке, образованной на искусственных поверхностях, бактерии обычно лишены капсул. Однако в культуре трансформированного бронхиального эпителия человека выявлены капсульные формы менингококка в толще биопленки [10]. Пока трудно сказать, какая из моделей – на искусственной поверхности или на бронхиальном эпителии – больше соответствует естественным условиям этого микроорганизма. У открепляющихся планктонных клеток, предназначенных для выхода в воздушную среду для передачи новому лицу, капсула имеется. Так формируется носительство, длящееся порой несколько недель и даже месяцев. Внутри биопленки менингококки защищены от многих антибиотиков, например, от пенициллина, чувствительность к которому у неразмножающихся клеток резко снижена. В то же время сохраняется чувствительность к рифампину [5], что вполне согласуется с различной эффективностью этих антибиотиков в отношении носительства. Плохо проникают в биопленки антитела и другие иммунологические факторы. Именно в биопленках происходит обмен генетическим материалом и иные события, приводящие к формированиию новых генетических, а значит и антигенных разновидностей. Таковы современные представления о возникновении и природе менингококкового носительства, длящегося порой от нескольких дней до недель и даже месяцев [10–13]. В случае длительного носительства патогенного или условно-патогенного микроорганизма говорят о персистенции [14].
Рассмотрим второй вариант – инвазию менингококка в организм человека. При этом отмечается скудная колонизация, слабое соединение друг с другом микроколоний, формирование тонкой одно-двухслойной неструктурированной биопленки. Происходит внедрение менингококка в эпителий, дальнейшее прохождение сквозь слизистую, и в ряде случаев – проникновение в кровь, на мозговые оболочки.
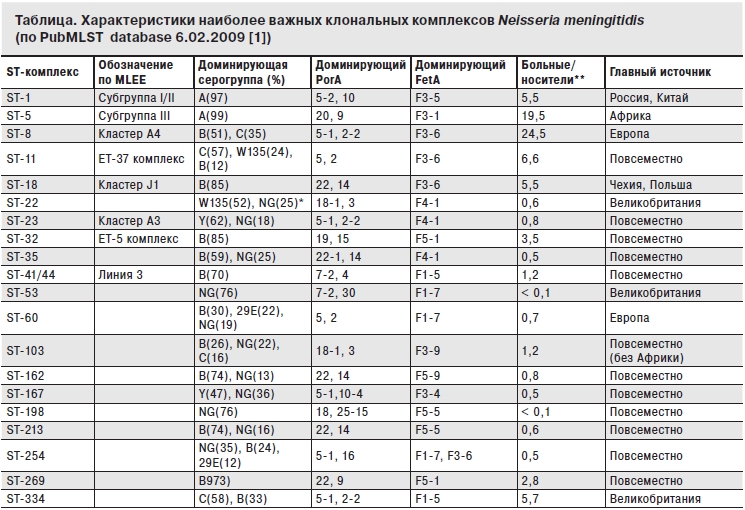
Такое двоякое поведение менингококка вызвано генетической неоднородностью его популяции. Благодаря внедрению метода мультилокусного энзимного электрофореза (MLEE) в 80-х гг., а затем – более современного мультилокусного секвенирования-типирования (MLST) среди менингококков выявлены неодинаковые по вирулентности генетические клоны. Генерализованные формы инфекции вызывают именно те клоны, которые слабо колонизируют и почти не образуют биопленку [1, 10, 15]. Их называют гиперинвазивными клонами, они высеваются от больных при вспышках и эпидемиях [16]. К 2009 г. в Генбанке было накоплено свыше 10 000 диких штаммов, представляющих 6751 генотип (сиквенс-тип – ST), которые по степени родства были объединены в 37 клональных комплексов (ST-комплексы). При этом принадлежность штамма к тому или иному генотипу не всегда совпадает с его антигенными свойствами. В таблице представлены 20 наиболее распространенных к 2009 г. клональных ST-комплексов в сопоставлении с их способностью вызывать преимущественно болезнь или носительство (по числу хранящихся штаммов), а также с их антигенной специфичностью. Кстати, штаммы клональных комплексов ST-1, SТ-41/44, ST-53, а также неизвестные ранее ST выявлены и на территории Москвы [17]. В популяциях микроорганизма постоянно возникают новые клоны с помощью генетических событий – трансформации, рекомбинаций, мутаций, трансдукции [8, 13, 15, 18]. Некоторые из них, благодаря наличию факторов патогенности и новых для населения антигенов, приобретают селективное преимущество перед циркулирующими ранее. Через несколько лет часть ST исчезнет из подобных таблиц, но зато добавятся новые, непредвиденные пока клоны. Необычайная генетическая пластичность возбудителей ОГМ (не только менингококка) позволяет им длительно циркулировать среди населения в различных формах взаимодействия с человеком, постоянно обновляя свою популяцию.
*NG – негруппируемый.
** Отношение вычислено по числу находящихся в PubMLST database изолятов, представленных членами научных коллективов по изучению нессерий.
На основании способности к инвазии или к образованию солидных биопленок M. Lappann и соавт. [11] предложили разделить генотипы менингококка на «распространителей» (англ. spreader) и «поселенцев» (англ. settler), причем к первым отнесены именно гиперинвазивные клоны. Естественно, что эти две группы не имеют четкой границы. Инвазивные штаммы-клоны не тратят энергию на формирование прочной биопленки, а стремятся проникнуть во внутреннюю среду организма. Считается, что отсутствие массивной колонизации компенсируется у этих штаммов высокой инвазивной активностью, чему способствует капсула, защищающая менингококк во внешней среде. Низкая способность этих штаммов к пленкообразованию частично объясняется отсутствием у них ряда аутолитических ферментов, приводящих к распаду бактериальных клеток и высвобождению ДНК, являющейся, как уже говорилось, фактором прикрепления биопленки к слою эпителия [9].
После закрепления и образования микроколоний пили начинают мешать тесному контакту бактерии с эпителиальной клеткой, необходимому для дальнейшей инвазии, и менингококк ретрагирует (втягивает) их. В ретракции пилей участвует структурный белок пилина PilT. На этом этапе клетка менингококка плотно прилегает к поверхностной мембране эпителиальной клетки. Теперь, в отсутствие пилей, роль адгезинов выполняют различные поверхностные белковые структуры клеточной стенки менингококка (рис. 2, см. на вклейке), главными из которых являются белки Ора и Орс, а также множество минорных белков NadA, NhhA, App, MspA, Tps (HrpA) и др. [3, 13, 19]. На этом этапе действуют межмолекулярные сигналы между бактериями и эпителиальными клетками, а также между бактериями. Плотное соприкосновение стенки бактерии с мембраной эпителиальной клетки позволяет возбудителю активно использовать ее межмолекулярные связи. Под влиянием этого вмешательства на поверхности эпителиальной клетки появляются выросты, обволакивающие микроб [13]. На этой стадии возможно его дальнейшее проникновение в клетку человека [1, 20]. Главную роль при этом играет белок клеточной стенки инвазин Орс. Наблюдения in vivo и ex vivo (на монослое бронхиального эпителия) T.C. Southerland и соавт. [21] показали, что проникновение менингококка в клетку (эндоцитоз) и прохождение сквозь нее (трансцитоз) происходит без повреждения клеточного слоя. Через 8 часов после колонизации под эпителием уже видны отдельные бактерии. На трансцитоз менингококка сквозь эпителий уходят примерно 1 сут. Нейссерии внедряются в эпителиальные клетки, находясь на «пъедесталах» – выростах с углублениями, что является результатом перестройки актинового скелета клетки человека под влиянием «обманных» межмолекулярных сигналов, исходящих от возбудителей. При этом связи между клетками эпителия остаются нерушимыми. Под микроколонией клетка хозяина образует так называемую кортикальную «бляшку», в которой и формируются сигналы по захвату паразита – эндоцитозу.
В период трансцитоза (прохождения сквозь клетку) менингококк снова экспрессирует капсулу [20, 21]. Капсула защищает находящиеся внутри клеток бактерии от дефенсинов – катионных антимикробных пептидов, факторов врожденного иммунитета. Прошедшие сквозь эпителий менингококки уже имеют капсулу. Прохождению сквозь эпителий и выходу из него способствуют находящиеся в эпителиальной клетке структуры – микротрубочки, образующие сеть [21].
В подслизистом слое менингококки встречаются с факторами иммунной защиты, в результате чего они или погибают, или вызывают местный воспалительный процесс. Это воспаление подслизистого пространства носоглотки соответствует клинической форме инфекции – назофарингиту.
Далее возбудитель проходит сквозь эндотелий мелких сосудов в кровяное русло (см. рис. 1, на вклейке), также используя адгезию к эндотелиальным клеткам (снаружи!), и белок Орс, применяя тактику «обманных» межмолекулярных сигналов. В подавляющем числе случаев проникшие в кровь N. meningitidis погибают под влиянием факторов врожденного и приобретенного иммунитета. Однако гиперинвазивным штаммам удается избежать этого; происходит размножение возбудителя в крови (бактериемия), нередко переходящая в сепсис и/или менингит. На этой стадии главным фактором патогенности является липоолигосахарад (ЛОС) клеточной стенки – эндотоксин. Частички ЛОС отрываются от поверхности бактерии в виде «пузырей». Именно ЛОС вызывает системную воспалительную реакцию, активирует комплемент и индуцирует выход каскада провоспалительных цитокинов – ИЛ-6, TNF-α, хемокинов, а также окиси азота. Еще в 1989 г. была показана прямая связь между уровнем ЛОС в кровотоке и летальностью [22]. Однако чтобы оказывать токсическое действие, паразит должен выжить в крови, богатой антибактериальными факторами. Для защиты от комплемента, антител и фагоцитоза у менингококка значительно утолщается капсула. Кроме того, менингококк активно связывает фактор Н (fH) благодаря наличию в клеточной оболочке специального липопротеина fHbp и таким способом защищает себя от бактерицидного действия сыворотки [23]. Другой поверхностный белок менингококка – PorA – связывает компонент комплемента С4 [3]. Устойчивости к сыворотке способствует и поверхностный белок инвазин Орс [19].
Как уже указывалось, находящиеся в кровотоке микроорганизмы получают возможность проникновения между мозговыми оболочками (pia mater и arachnoidea mater) в спинномозговую жидкость (СМЖ). СМЖ является благоприятной средой для размножения патогенных бактерий, так как в норме лишена и комплемента, и антител, а в отсутствие последних даже массовый приток нейтрофилов мало эффективен. Однако ликвор надежно защищен так называемым гематоэнцефалическим барьером (ГЭБ) от микроорганизмов, находящихся в кровотоке. ГЭБ состоит из прочно скрепленного эндотелия мозговых капилляров и различных фагоцитирующих периваскулярных клеток (перициты, макрофаги, клетки микроглии), а также астроцитов, что в совокупности формирует иммунологический барьер [4, 24].
Несмотря на наличие ГЭБ, в настоящее время господствует теория гематогенного пути инфицирования мозговых оболочек. Однако еще в середине ХХ века считалось, что менингококк попадает на мозговые оболочки со слизистой носа по обонятельным нервам, но вскоре гематогенная теория взяла верх. В 2010 г. шведские ученые [25] показали такую возможность в эксперименте на трансгенных мышах, экспрессирующих один из рецепторов для адгезинов – человеческий белок CD46, при интраназальном заражении гиперинвазивным клоном менингококка (ST-11). Они поддерживают свое заключение на основании клинически и бактериологически подтвержденных случаев менингита у людей без признаков бактериемии и считают, что этот путь может существовать наряду с гематогенным. Проникновение возбудителей в СМЖ по обонятельным нервам выявлено для пневмококка и ряда вирусов [26].
Но вернемся к общепризнанному гематогенному пути. Как показали недавние исследования [27, 28], менингококки образуют микроколонии на внутренней поверхности эндотелия, как и на эпителии, что позволяет им удерживаться и не быть смытыми кровотоком. Однако бактерии внедряются в толщу эндотелиальной клетки гораздо глубже, чем в эпителиальную. В этом процессе опять же участвуют пили, причем один их структурных белков – пилин PilV – ответственен за образование именно глубоких вдавлений или ячеек на эндотелии для укрепления менингококковых клеток. Эти ячейки образуются благодаря выростам эндотелия, индуцированным менингококком путем вмешательства в межмолекулярные сигналы, сходно с процессом, который наблюдается при взаимодействии с эпителием. Несмотря на сходство, пути межмолекулярной сигнализации при этих процессах различаются [29]. Образование укрепленных микроколоний на эндотелии мозговых капилляров, возможно, является аналогом формирования биопленок на эпителии.
На рубеже ХХ и ХХI веков была показана инвазия менингококка в эндотелий благодаря белку инвазину Орс и его связыванию с белками человека – фибронектином и соответствующим ему интегриновым рецептором [19] путем трансцитоза. Казалось, что вопрос о проходе менингококка из кровотока в СМЖ решен [30]. Однако недавно было показано, что основной путь преодоления менингококком стенки мозгового капилляра – парацеллюлярный, т. е. между клетками эндотелия мозговых сосудов [4]. Именно мозговые капилляры имеют очень прочные межклеточные соединения, отличные от таковых у других капилляров тела человека. Эти соединения состоят из трансмембранных белков, образующих чередующиеся плотные и липкие связи, которые в свою очередь взаимосвязаны с актиновым скелетом и сигнальными молекулами внутри эндотелиальных клеток. После формирования микроколонии под ней на клетке эндотелия формируется «бляшка», под которой концентрируются сигнальные молекулы и трансмембранные белки. Молекулярный механизм образования выростов, кортикальной бляшки и последующих событий изучен достаточно полно [24]. Вмешательство менингококка во внутриклеточные сигналы приводит не только и не столько к его поглощению и трансцитозу сквозь эндотелий, сколько к разрыву межклеточных связей в стенках мозговых капилляров. В разрывы между эндотелиальными клетками устремляется менингококк и оказывается в субарахноидальном пространстве. Продукты распада менингококка (ЛОС, пептидогликан) способствуют выработке провоспалительных цитокинов, привлекающих в свою очередь в СМЖ огромное количество фагоцитов – нейтофилов и макрофагов. Так формируется гнойное воспаление мозговых оболочек – гнойный менингит.
По сходной схеме проходят путь и другие возбудители ОГМ – внеклеточные комменсалы слизистых оболочек, хотя у каждого вида имеются свои отличия.
Пневмококк, находящийся на слизистой оболочке дыхательных путей, с помощью ферментов (нейраминидаза, IgA-протеаза и др.) преодолевает слой слизи, воздействие секреторных антител, биение ресничек (благодаря пневмолизину) и адгезируется на поверхности эпителия. Капсульный слой на стадии прикрепления почти отсутствует. Адгезия осуществляется с помощью поверхностных холин-связывающих белков CbpA (он же PspC) и PspA, недавно открытого, связанного с пилями белка RrgA [31], а также поверхностных липотейхоевых (ЛТК) и тейхоевых кислот (ТК). Подобно менингококку, пневмококк путем вмешательства в сигнальные пути внутри эпителиальной клетки вынуждает последнюю фагоцитировать себя (эндоцитоз) и проходить сквозь нее (трансцитоз). Трансцитозу пневмококка способствует наличие воспалительного процесса слизистой (вирусная инфекция, синусит, отит и т. п.). Но в отличие от менингококка уже на этом этапе пневмококк применяет и парацеллюлярный проход – между клетками эпителия. Для этого паразит использует плазминоген крови человека, фиксируя его на своей поверхности и превращая его в плазмин – мощную протеазу, разрушающую межклеточные связи [32]. Субэпителиальное соединительнотканное пространство преодолевается благодаря ферменту гиалуронидазе (гиалуронан-лиаза) [33]. На этой стадии паразиту уже необходима капсула. Подобно менингококку, пневмококк проходит сквозь эндотелий сосудов субэпителиального пространства путем трансцитоза, используя свой адгезивный сигнальный белок PspC, и попадает в кровоток. В крови пневмококк защищается от фагоцитоза благодаря капсуле, от бактерицидного действия сыворотки – связывая фактор Н и компоненты комплемента. Он также адгезируется на внутренней поверхности мозговых капилляров, разрушает с помощью пневмолизина (пневмококкового токсина) межклеточные связи эндотелия и проникает в пространство между мягкой и паутинной мозговой оболочкой, т. е. в СМЖ. Обильно размножающиеся в СМЖ пневмококки быстро входят в стационарную фазу роста и ликвор наполняется продуктами распада его клеток. Продукты распада клеточной стенки пневмококка (преимущественно ЛТК и ТК) являются индукторами провоспалительных цитокинов. Возникает обильный приток в СМЖ лейкоцитов, в основном нейтрофилов, которые оседают на мозговых оболочках в виде макроскопических скоплений. Эти же бактериальные продукты повреждают мозговую кору [32]. Распаду бактериальных клеток содействуют также бактерицидные антибиотики (например, цефтриаксон), применяющиеся для лечения ОГМ. В связи с индукцией чрезмерной воспалительной реакции продуктами распада пневмококка высказываются предложения применять только бактериостатические антибиотики, например, даптомицин, рифампин, моксифлоксацин [32].
Давно известно, что далеко не все разновидности пневмококка способны вызывать менингит. Из 95 известных ныне серотипов лишь не более 30 выделяются при ОГМ и бактериемиях. Частота их выявления на разных территориях и в разные периоды времени неодинакова. При этом внутри серотипов также найдены гиперинвазивные генетические клоны, не всегда совпадающие с принадлежностью к определенному сероварианту, как это имеет место у менингококков.
Недавно показано, что гиперинвазивный штамм стрептококка серогруппы В (СГВ) проходит через носоглоточный эпителий человека между клетками [34], но при переходе из кровотока в СМЖ использует трансцитоз. С помощью липких молекул (PilA) на конце пилей стрептококки прикрепляются к коллагену – матричному экстрацеллюлярному белку, находящемуся вблизи сосудов. Коллаген в свою очередь соединяется с интегрином α2β1, расположенным на поверхности эндотелия мозгового сосуда. После прикрепления интегрин посылает клетке сигналы по увеличению выработки провоспалительных цитокинов и хемокина ИЛ-8, вызывающих массовый приток нейтрофилов. Нейтрофилы, прилипающие к эндотелиальным клеткам, повышают их проницаемость, что способствует инвазии СГВ [35]. Преодолевать защитные барьеры организма также способны лишь отдельные генетические разновидности стрептококков группы В. Так, наиболее часто инвазивные формы вызывают серотипы Ia, Ic, IIc, III и V, а внутри серотипов описаны генетические клоны, характерные преимущественно для носительства или для острого менингита [36].
Открытие инвазивных клонов в популяциях видов возбудителей ОГМ существенно повлияло на представления о специфической профилактике этих заболеваний. Так. M.C.J. Maiden и соавт. [37] считают, что здравоохранение должно контролировать (в смысле «ограничивать») менингококковую инфекцию путем элиминации из циркулирующей популяции только капсульных гиперинвазивных клонов. Им удалось достичь этой цели благодаря полисахаридным конъюгированным вакцинам против соответствующих штаммoв в Англии [38] и в Нидерландах. В связи с этим, по нашему мнению, мониторинг клональной структуры циркулирующих возбудителей ОГМ должен стать неотьемлимой частью эпидемиологического надзора при этих заболеваниях.



{kind=link}